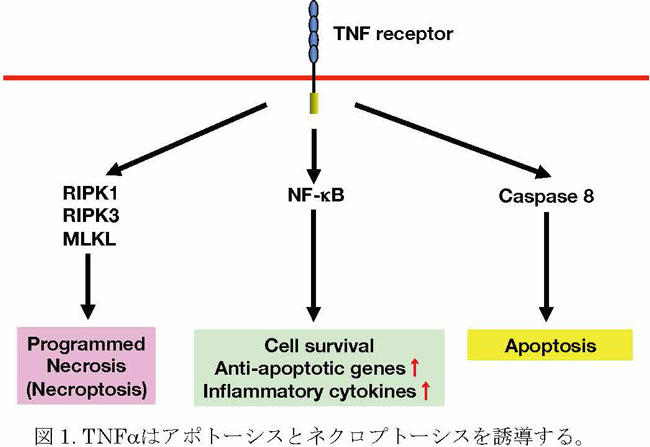
理事⻑就任にあたり2020年06月19日
細胞死との3度の出会い
太⽥先⽣、三浦先⽣と偉⼤な先⽣⽅が理事⻑をされた後で、甚だ実⼒不⾜ですが、この7⽉から理事⻑を務めさせていただくことになりました東邦⼤学医学部⽣化学講座の中野でございます。
私と細胞死との初めての出会いは、⼤学院時代に遡ります。医者にはなったものの臨床医としての限界を感じ、1991 年に私は⼤学院に⼊ることを決意しました。⼤学院⽣として選んだ研究室はT 細胞のシグナル伝達の研究をしていた斎藤隆教授が主宰されており、全く細胞死とは関係のない研究室でした。しかしそこに⽶国のDNAX 研究所から新井賢⼀先⽣の研究室出⾝の宮武先⽣(現⿇布⼤学教授)が助教として赴任され、T 細胞ハイブリドーマで⾒られるactivation-induced cell death (AICD)と細胞周期との関連のプロジェクトを⽴ち上げられ、そのグループ(といってもスタートは2 ⼈だけ)に私が参加したのが細胞死との最初の出会いでした。その後1995 年にLa Jolla Institute for Allergy and ImmunologyにいたDouglas Green らがactivation-induced cell death の本体はTCR 刺激によりFasL が誘導され細胞⾃律的にアポトーシスに陥るという論⽂をNature に出し、結果的には細胞周期とactivation-induced cell death は直接は関係ないという結論になりました。その論⽂が出る前(1993 年)に⻑⽥研にいた須⽥先⽣(現⾦沢⼤学教授)によりFasL のクローニングがなされました。ただ、当時はアポトーシス研究の黎明期であり、哺乳類の最初のカスパーゼであるICE がYuan のラボにいた三浦先⽣によりクローニングされたのが1993 年で、アポトーシス実⾏のメカニズムの⼤部分はブラックボックスでした。当然ゲノムプロジェクトも始まっておらずマウスやヒトで全てのタンパク質をコードする遺伝⼦が何万個あるかもわかっておらず(⼀時は10 万個と⾔われていたこともありました)、かつEST のデータベースさえなかったような時代です。このような時代(1992 年)に本庶先⽣の研究室の⽯⽥靖雄先⽣(現奈良先端技術⼤学准教授)がT cell hybridoma をTCR で刺激してアポトーシスを誘導した時の細胞と、未刺激の細胞からRNA を調製してsubtraction library を作成し、アポトーシスに関係しているだろう遺伝⼦として同定された遺伝⼦がPD (Program cellDeath)-1 でした。今から思えばPD-1 は活性化したT 細胞で発現する遺伝⼦だったわけで す。PD-1 が今のような状況になるとは、その当時は誰も思っていなかったと思います。
なんとか別の仕事をまとめて学位論⽂として発表し、その後1995 年に順天堂⼤学医学部の奥村研究室の助教として赴任しました。赴任後はTRAF5 という遺伝⼦をクローニングし、TNF 受容体を介するシグナル伝達やNF-kB の活性化のメカニズムの研究をしていたのですが、John Reed 研に留学してその後⽇本に帰国された現京⼤教授の⾼橋良輔先⽣(⾼橋先⽣とは⽶国留学前から共通の知⼈を介して⾯識がありました)から1999 年の⽣化学会の細胞死関連のシンポジウムのシンポジストとして招待していただきました。細胞死のシンポジウムで発表させていただく機会を与えていただいたことで、私が細胞死研究を強く意識した2回⽬(2 度⽬の細胞死との出会い)になりました。
その後2001 年に当時医科⻭科⼤学の教授になられていた⼀條先⽣(現東京⼤学教授)から誘われて岡崎の⽣理研の岡⽥泰伸先⽣が毎年世話役として開催されていた「細胞死研究会」に誘っていただいたのが、細胞死との3 度⽬の、かつ私のその後の研究の⽅向性を決めた決定的な出会いになりました。「細胞死研究会」では⽇本の細胞死研究の中核を担っていた⻑⽥重⼀先⽣、辻本賀英先⽣、⽶原伸先⽣、鍔⽥武志先⽣、⾼橋良輔先⽣、⼀條秀憲先⽣、垣塚彰先⽣、三浦正幸先⽣、清⽔重⾂先⽣、仁科博史先⽣、後藤由季⼦先⽣らが参加されておりました。これらの⽅々の前で毎年30 分間⾃分の1年間の研究成果を発表することは胃が痛くなるようなプレッシャーでした。しかしこの研究会は私の⽇本における細胞死研究関連の⼈々との⼈脈を形成する上で⼤きな糧となりましたが、残念ながらこの研究会は岡⽥先⽣が⽣理研の所⻑に就任(2007 年)されご多忙になられたということで2009 年に消滅しました。
⼀⽅1992 年に⼭⽥武先⽣たちが中⼼となって発⾜したアポトーシス研究会は、1997 年に東海アポトーシス研究会と合併し、2009 年に太⽥先⽣の強いリーダーシップのも と⽇本Cell Death 学会へと昇格しました。岡崎の細胞死研究会に参加されていた⼈たちも、Cell Death 学会に参加することになり、現在に⾄っております。⼀⽅で、細胞死研究の新たなハブとなるべく細胞死に関する新学術領域の⽴ち上げは、毎年のように不採択となりながらも地道に継続され、⽥中正⼈先⽣のもとでようやく2014 年に新学術領域「ダイイングコード」が⽴ち上がり、私も微⼒ながら貢献させていただきました。残念ながらこの領域も2019 年3⽉で終了してしまい、現時点で細胞死関連の⼈々が参加し、討論できる場は⽇本Cell Death 学会のみになってしまいました。今後はこの学会をさらに発展させ、若⼿の細胞死研究者の育成や、広く研究者の⼈脈を広げる場としても盛り上げていきたいと思っておりますので、よろしくお願いいたします(⽇本Cell Death 学会Newsletter Vol. 25)。